LY96 Antibody
-
中文名稱:LY96兔多克隆抗體
-
貨號(hào):CSB-PA049918
-
規(guī)格:¥1100
-
圖片:
-
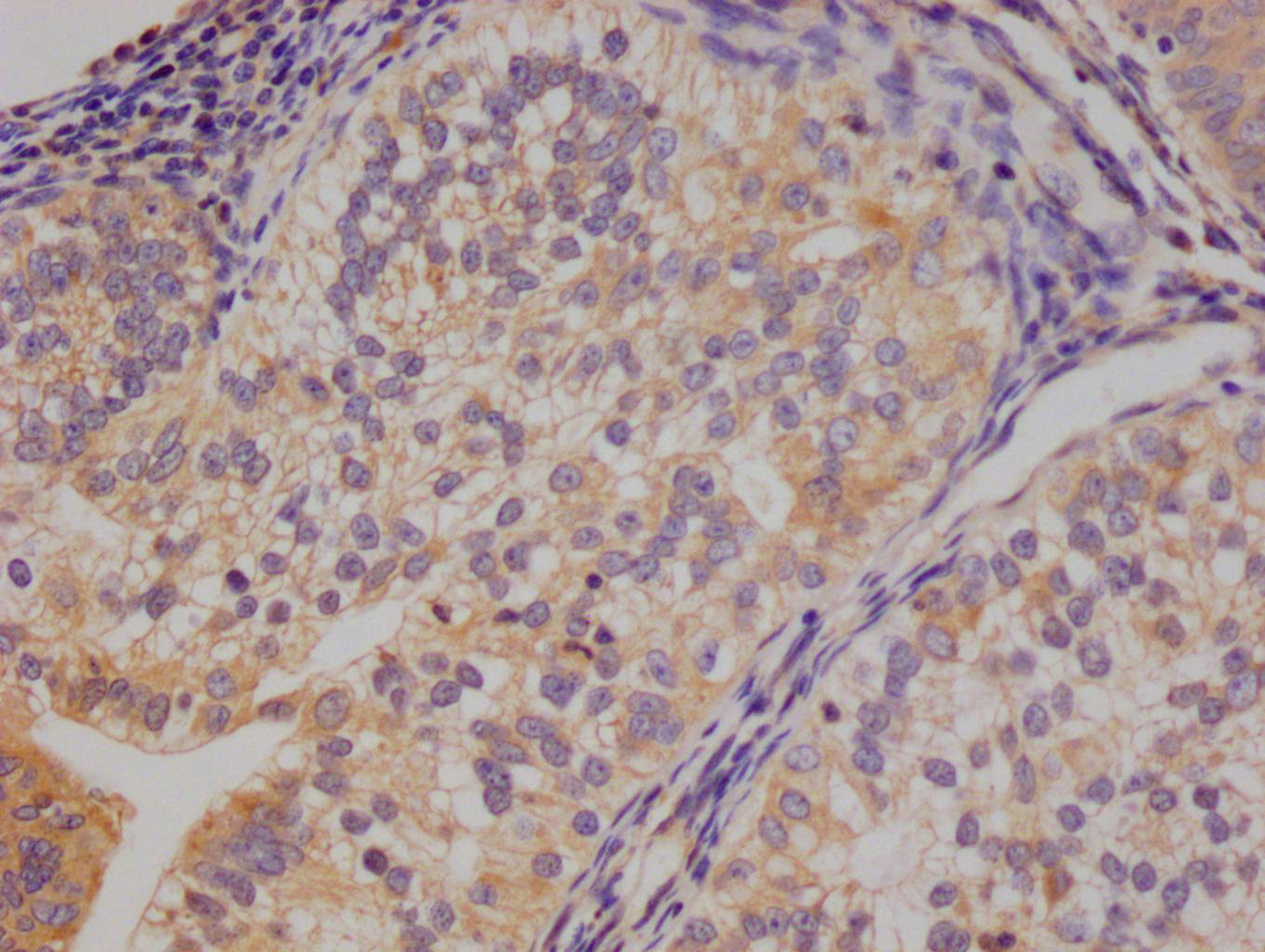
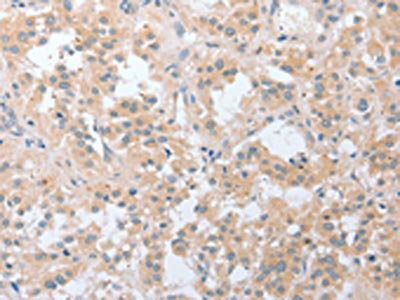 The image on the left is immunohistochemistry of paraffin-embedded Human thyroid cancer tissue using CSB-PA049918(LY96 Antibody) at dilution 1/20, on the right is treated with fusion protein. (Original magnification: ×200)
The image on the left is immunohistochemistry of paraffin-embedded Human thyroid cancer tissue using CSB-PA049918(LY96 Antibody) at dilution 1/20, on the right is treated with fusion protein. (Original magnification: ×200) -
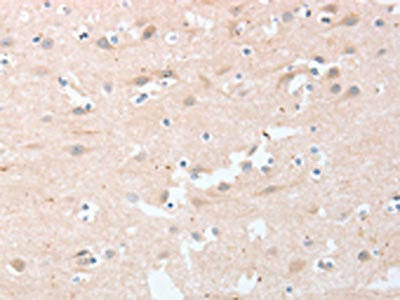 The image on the left is immunohistochemistry of paraffin-embedded Human brain tissue using CSB-PA049918(LY96 Antibody) at dilution 1/20, on the right is treated with fusion protein. (Original magnification: ×200)
The image on the left is immunohistochemistry of paraffin-embedded Human brain tissue using CSB-PA049918(LY96 Antibody) at dilution 1/20, on the right is treated with fusion protein. (Original magnification: ×200) -
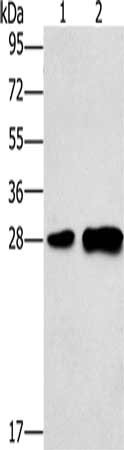 Gel: 10%SDS-PAGE, Lysate: 40 μg, Lane 1-2: Mouse liver tissue, 231 cells, Primary antibody: CSB-PA049918(LY96 Antibody) at dilution 1/500, Secondary antibody: Goat anti rabbit IgG at 1/8000 dilution, Exposure time: 1 minute
Gel: 10%SDS-PAGE, Lysate: 40 μg, Lane 1-2: Mouse liver tissue, 231 cells, Primary antibody: CSB-PA049918(LY96 Antibody) at dilution 1/500, Secondary antibody: Goat anti rabbit IgG at 1/8000 dilution, Exposure time: 1 minute
-
-
其他:
產(chǎn)品詳情
-
Uniprot No.:
-
基因名:
-
別名:ESOP 1 antibody; ESOP-1 antibody; ESOP1 antibody; LY 96 antibody; Ly-96 antibody; LY96 antibody; LY96_HUMAN antibody; Lymphocyte antigen 96 antibody; md 2 antibody; MD 2 protein antibody; MD2 protein antibody; Myeloid differentiation protein 2 antibody; Protein MD 2 antibody; Protein MD-2 antibody; Protein MD2 antibody
-
宿主:Rabbit
-
反應(yīng)種屬:Human,Mouse
-
免疫原:Fusion protein of Human LY96
-
免疫原種屬:Homo sapiens (Human)
-
標(biāo)記方式:Non-conjugated
-
抗體亞型:IgG
-
純化方式:Antigen affinity purification
-
濃度:It differs from different batches. Please contact us to confirm it.
-
保存緩沖液:-20°C, pH7.4 PBS, 0.05% NaN3, 40% Glycerol
-
產(chǎn)品提供形式:Liquid
-
應(yīng)用范圍:ELISA,WB,IHC
-
推薦稀釋比:
Application Recommended Dilution ELISA 1:1000-1:2000 WB 1:500-1:2000 IHC 1:25-1:100 -
Protocols:
-
儲(chǔ)存條件:Upon receipt, store at -20°C or -80°C. Avoid repeated freeze.
-
貨期:Basically, we can dispatch the products out in 1-3 working days after receiving your orders. Delivery time maybe differs from different purchasing way or location, please kindly consult your local distributors for specific delivery time.
產(chǎn)品評(píng)價(jià)

相關(guān)產(chǎn)品
靶點(diǎn)詳情
-
功能:Binds bacterial lipopolysaccharide (LPS). Cooperates with TLR4 in the innate immune response to bacterial lipopolysaccharide (LPS), and with TLR2 in the response to cell wall components from Gram-positive and Gram-negative bacteria. Enhances TLR4-dependent activation of NF-kappa-B. Cells expressing both LY96 and TLR4, but not TLR4 alone, respond to LPS.
-
基因功能參考文獻(xiàn):
- sTLR4/MD-2 complex inhibits colorectal cancer migration and invasiveness in vitro and in vivo by lncRNA H19 down-regulation. PMID: 29036538
- MD-2 siRNA or plasmid further confirmed the efficacy of Iturin A by suppressing MD-2/TLR4 signaling pathway. The in silico docking study showed that the Iturin A interacted well with the MD-2 in MD-2/TLR4 receptor complex. Conclusively, inhibition of MD-2/TLR4 complex with Iturin A offered strategic advancement in cancer therapy. PMID: 28347875
- data confirm that engineered human cells expressing TLR4, MD2 and CD14 can respond to CMP with NF-kappaB activation and the response can be influenced by variations in CMP-mannosylation PMID: 29281684
- The observations suggest that MD-2 helps to regulate lipopolysaccharide-induced NLRP3 inflammasome activation and the inflammatory response in NR8383 cells, and likely does so by affecting MyD88/NF-kappaB signaling. PMID: 28654770
- MD2 plays an important role in induction of allergic sensitization to cat dander and common pollens relevant to human allergic diseases. PMID: 26586036
- we report that exogenous CnB is taken up by cells in a time- and concentration-dependent manner via clathrin-dependent receptor-mediated internalization. Our findings further confirm that uptake is mediated by the TLR4/MD2 complex together with the co-receptor CD14 PMID: 27090571
- In this study, a novel naturally occurring spliceosome of human MD2, termed as MD2-T3, has been identified. PMID: 27317890
- Results show that cigarette smoke may alter innate immune responses reducing the expression of the MD2, a molecule with an important role in TLR4 signaling. PMID: 26068048
- Predominantly hydrophobic interactions between MD-2 and TLR4 contribute to the stabilization of the TLR4/MD-2/metal ion complex in a conformation that enables activation. PMID: 25803856
- The intensity of the intra-amniotic inflammatory response to bacteria or perhaps to other TLR4 ligands may be facilitated through synthesis and release of sMD2 by the amniochorion. PMID: 25605324
- Three genes (LY96, IL8 DPR) were significantly downregulated over time. This finding was confirmed in a validation cohort of stroke patients (n=8). PMID: 25124890
- The study revealed the impact of specific residues and regions of MD-2 on the binding of lipolysaccharides and TLR4. PMID: 26320630
- Gene polymorphisms of MD2 and GM2A were associated with the occurrence or severity of neonatal necrotizing enterocolitis. PMID: 25816011
- In patients undergoing CABG surgery, we found genetic polymorphisms in LY96 associated with decreased risk of postoperative AF. PMID: 26385043
- Aminoarabinose residues in lipid A contribute to Burkholderia lipid A binding to the TLR4.Myeloid Differentiation Factor 2 complex. PMID: 26160169
- TLR4 along with its accessory protein MD-2 builds a heterodimeric complex that specifically recognizes lipopolysaccharides, which are present on the cell wall of gram-negative bacteria, activating the immune response. (Review) PMID: 25515751
- genetic polymorphism is associated with asthma exacerbations in children PMID: 24902762
- In monocytes of sepsis, MD-2 expression was found to be regulated by STAT1. PMID: 24858600
- Substitution of hMD-2 residue V82 with an amino acid residue with a bulkier hydrophobic side chain enables activation of TLR4/MD-2. PMID: 25203747
- This study suggests that genetic variants of LY96 may modulate the effect of smoking on carotid plaque burden. PMID: 24954085
- overexpression of MD-2s led to marked reduction in markers of tissue injury and inflammation in models of lung inflammation PMID: 25576596
- Purified monomeric ligand.MD-2 complexes reveal molecular and structural requirements for activation and antagonism of TLR4 by Gram-negative bacterial endotoxins. PMID: 24895101
- Studies suggest that myeloid differentiation 2 (MD-2) as a therapeutic target for sepsis. PMID: 24896325
- Mechanistically, engagement of MD-2 by PTX3-opsonized Aspergillus conidia activated the TLR4/Toll/IL-1R domain-containing adapter inducing IFN-beta-dependent signaling pathway converging on IL-10. PMID: 25049357
- Collectively, these data showed for the first time that, HIV-1 Tat interacts physically with high affinity with TLR4-MD2 to promote proinflammatory cytokines (TNF-alpha) and the immunosuppressive cytokine IL-10 both. PMID: 24165011
- Data suggest that zhankuic acid A (ZAA), which competes with LPS for binding to MD-2 as a TLR4/MD-2 antagonist, may be a potential therapeutic agent for gram-negative bacterial infections. PMID: 24532584
- these findings indicate that MD-2 is involved in IFN-gamma signalling and IFN-gamma-augmented MMP-1 up-regulation by LPS. PMID: 23800176
- Extracellular complex formation of TLR4 with secreted MD-2 was detectable using monoclonal antibodies. PMID: 24021278
- both oral and enteric C. concisus strains upregulated expression of TLR4 and MD-2 in HT-29 cells. PMID: 23437263
- Consistently, soluble TLR4 expressed without MD2 inhibited metal- but not LPS-induced responses, opening new therapeutic perspe PMID: 23059983
- Respiratory syncytial virus F protein requires MD-2 for the induction of the TLR4-mediated inflammatory response. PMID: 22872782
- In LOS.MD-2 complexes, one of the six fatty acyl chains of LOS is more susceptible to paramagnetic attenuation, suggesting protrusion of that fatty acyl chain from the hydrophobic pocket of MD-2, independent of association with TLR4. PMID: 22433852
- Lipopolysaccharide cytokine response was efficiently and equally abolished by MD-2 and CD14 neutralization; the response induced by whole E. coli bacteria was only modestly reduced by MD-2 neutralization, whereas CD14 neutralization was more efficient. PMID: 21948372
- have identified specific structural and dynamic features of the MD-2-endotoxin complexes that may control dimerization of TLR4 molecules. PMID: 21924775
- The association between occupational endotoxin exposure and wheeze in agricultural workers was significantly modified by genetic variants in CD14 and MD2 PMID: 21389010
- Data demonstrate that during bacterial infection, newborns amplify the TLR2-MyD88 pathway in G+ bacterial infection and the TLR4/MD2/MyD88 pathway in G- bacterial infection, implicating the innate immune pathway in response to bacterial infection. PMID: 20805788
- higher expression levels in tissue from patients with colorectal cancer PMID: 20699280
- Computational study of the interactions of the unmodified dendrimer, glucosamine, and of the partially glycosylated dendrimer with TLR4 and MD-2 using molecular docking and molecular dynamics techniques. PMID: 21738462
- tyrosine phosphorylation of MD-2 is important for signaling following exposure to lipopolysaccharide PMID: 21918188
- Cytokines can enhance TLR4 and MD-2 expressions and promote the reaction with LPS in intestinal epithelial cells. PMID: 20848773
- Interferon-gamma-induced MD-2 protein expression and lipopolysaccharide (LPS) responsiveness in corneal epithelial cells is mediated by Janus tyrosine kinase-2 activation and direct binding of STAT1 protein to the MD-2 promoter. PMID: 21572044
- In this study, we produced variants of MD-1 and MD-2 in Pichia pastoris. PMID: 21130168
- Innate immune recognition of meningococcal capsular polysaccharides by macrophages can occur via TLR2- and TLR4-MD-2 pathways. PMID: 21191086
- Neisserial lipooligosaccharides from different meningococcal and gonococcal strains have different potencies to activate NF-kappaappaB through the Toll-like receptor 4-MD-2 pathway in monocytes. PMID: 21037101
- Data conclude that these gene variants in the MD-2 gene promoter region are not associated with tuberculosis, and apparently do not play a role in susceptibility to tuberculosis in the Chinese population. PMID: 20730709
- species-specific activation of lipid IV(A) PMID: 20592019
- increased expression in intestinal epithelial cells in patients with inflammatory bowel disease PMID: 19710105
- In this study, we have identified a novel alternatively spliced isoform of human MD-2, termed MD-2 short (MD-2s), which lacks the region encoded by exon 2 of the MD-2 gene. PMID: 20435923
- Polymorphisms of the promoter region of MD-2 gene at position - 1625 C/G is correlated with MODS and sepsis after severe trauma in Chinese Han population. PMID: 19209789
- sMD-2 and sCD14 inhibit the growth of both Gram-positive and Gram-negative bacteria. PMID: 20377740
顯示更多
收起更多
-
亞細(xì)胞定位:Secreted, extracellular space. Secreted.
-
數(shù)據(jù)庫鏈接:
Most popular with customers
-
-
YWHAB Recombinant Monoclonal Antibody
Applications: ELISA, WB, IF, FC
Species Reactivity: Human, Mouse, Rat
-

Phospho-YAP1 (S127) Recombinant Monoclonal Antibody
Applications: ELISA, WB, IHC
Species Reactivity: Human
-

-

-

-

-