Recombinant Mouse Circadian locomoter output cycles protein kaput (Clock), partial
-
中文名稱:Recombinant Mouse Circadian locomoter output cycles protein kaput(Clock) ,partial,Yeast
-
貨號:CSB-YP005574MO
-
規(guī)格:
-
來源:Yeast
-
其他:
-
中文名稱:Recombinant Mouse Circadian locomoter output cycles protein kaput(Clock) ,partial,Yeast
-
貨號:CSB-EP005574MO
-
規(guī)格:
-
來源:E.coli
-
其他:
-
中文名稱:Recombinant Mouse Circadian locomoter output cycles protein kaput(Clock) ,partial,Yeast
-
貨號:CSB-EP005574MO-B
-
規(guī)格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:Recombinant Mouse Circadian locomoter output cycles protein kaput(Clock) ,partial,Yeast
-
貨號:CSB-BP005574MO
-
規(guī)格:
-
來源:Baculovirus
-
其他:
-
中文名稱:Recombinant Mouse Circadian locomoter output cycles protein kaput(Clock) ,partial,Yeast
-
貨號:CSB-MP005574MO
-
規(guī)格:
-
來源:Mammalian cell
-
其他:
產品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:Clock
-
Uniprot No.:
-
別名:ClockCircadian locomoter output cycles protein kaput; mCLOCK; EC 2.3.1.48
-
種屬:Mus musculus (Mouse)
-
蛋白長度:Partial
-
蛋白標簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產品評價

靶點詳情
-
功能:Transcriptional activator which forms a core component of the circadian clock. The circadian clock, an internal time-keeping system, regulates various physiological processes through the generation of approximately 24 hour circadian rhythms in gene expression, which are translated into rhythms in metabolism and behavior. It is derived from the Latin roots 'circa' (about) and 'diem' (day) and acts as an important regulator of a wide array of physiological functions including metabolism, sleep, body temperature, blood pressure, endocrine, immune, cardiovascular, and renal function. Consists of two major components: the central clock, residing in the suprachiasmatic nucleus (SCN) of the brain, and the peripheral clocks that are present in nearly every tissue and organ system. Both the central and peripheral clocks can be reset by environmental cues, also known as Zeitgebers (German for 'timegivers'). The predominant Zeitgeber for the central clock is light, which is sensed by retina and signals directly to the SCN. The central clock entrains the peripheral clocks through neuronal and hormonal signals, body temperature and feeding-related cues, aligning all clocks with the external light/dark cycle. Circadian rhythms allow an organism to achieve temporal homeostasis with its environment at the molecular level by regulating gene expression to create a peak of protein expression once every 24 hours to control when a particular physiological process is most active with respect to the solar day. Transcription and translation of core clock components (CLOCK, NPAS2, ARNTL/BMAL1, ARNTL2/BMAL2, PER1, PER2, PER3, CRY1 and CRY2) plays a critical role in rhythm generation, whereas delays imposed by post-translational modifications (PTMs) are important for determining the period (tau) of the rhythms (tau refers to the period of a rhythm and is the length, in time, of one complete cycle). A diurnal rhythm is synchronized with the day/night cycle, while the ultradian and infradian rhythms have a period shorter and longer than 24 hours, respectively. Disruptions in the circadian rhythms contribute to the pathology of cardiovascular diseases, cancer, metabolic syndromes and aging. A transcription/translation feedback loop (TTFL) forms the core of the molecular circadian clock mechanism. Transcription factors, CLOCK or NPAS2 and ARNTL/BMAL1 or ARNTL2/BMAL2, form the positive limb of the feedback loop, act in the form of a heterodimer and activate the transcription of core clock genes and clock-controlled genes (involved in key metabolic processes), harboring E-box elements (5'-CACGTG-3') within their promoters. The core clock genes: PER1/2/3 and CRY1/2 which are transcriptional repressors form the negative limb of the feedback loop and interact with the CLOCK|NPAS2-ARNTL/BMAL1|ARNTL2/BMAL2 heterodimer inhibiting its activity and thereby negatively regulating their own expression. This heterodimer also activates nuclear receptors NR1D1/2 and RORA/B/G, which form a second feedback loop and which activate and repress ARNTL/BMAL1 transcription, respectively. Regulates the circadian expression of ICAM1, VCAM1, CCL2, THPO and MPL and also acts as an enhancer of the transactivation potential of NF-kappaB. Plays an important role in the homeostatic regulation of sleep. The CLOCK-ARNTL/BMAL1 heterodimer regulates the circadian expression of SERPINE1/PAI1, VWF, B3, CCRN4L/NOC, NAMPT, DBP, MYOD1, PPARGC1A, PPARGC1B, SIRT1, GYS2, F7, NGFR, GNRHR, BHLHE40/DEC1, ATF4, MTA1, KLF10 and also genes implicated in glucose and lipid metabolism. Promotes rhythmic chromatin opening, regulating the DNA accessibility of other transcription factors. May play a role in spermatogenesis; contributes to the chromatoid body assembly and physiology. The CLOCK-ARNTL2/BMAL2 heterodimer activates the transcription of SERPINE1/PAI1 and BHLHE40/DEC1. The preferred binding motif for the CLOCK-ARNTL/BMAL1 heterodimer is 5'-CACGTGA-3', which contains a flanking Ala residue in addition to the canonical 6-nucleotide E-box sequence. CLOCK specifically binds to the half-site 5'-CAC-3', while ARNTL binds to the half-site 5'-GTGA-3'. The CLOCK-ARNTL/BMAL1 heterodimer also recognizes the non-canonical E-box motifs 5'-AACGTGA-3' and 5'-CATGTGA-3'. CLOCK has an intrinsic acetyltransferase activity, which enables circadian chromatin remodeling by acetylating histones and nonhistone proteins, including its own partner ARNTL/BMAL1. Represses glucocorticoid receptor NR3C1/GR-induced transcriptional activity by reducing the association of NR3C1/GR to glucocorticoid response elements (GREs) via the acetylation of multiple lysine residues located in its hinge region. The acetyltransferase activity of CLOCK is as important as its transcription activity in circadian control. Acetylates metabolic enzymes IMPDH2 and NDUFA9 in a circadian manner. Facilitated by BMAL1, rhythmically interacts and acetylates argininosuccinate synthase 1 (ASS1) leading to enzymatic inhibition of ASS1 as well as the circadian oscillation of arginine biosynthesis and subsequent ureagenesis. Drives the circadian rhythm of blood pressure through transcriptional activation of ATP1B1.
-
基因功能參考文獻:
- The expression of Epcam mRNA, which is a functional marker of potential hepatic stem-like cells, was controlled by LEF1, which was regulated by CLOCK. PMID: 29958886
- Results indicate that circadian locomotor output cycles kaput protein (CLOCK) overexpression triggers the formation of atherosclerotic plaques by directly upregulating plasminogen activator inhibitor 1 (PAI-1) expression. PMID: 30124738
- Low expression of CLOCK protein is associated with kidney tumor. PMID: 29271044
- CLOCK temporally gates mast cell responses to IL-33 via regulation of ST2 expression. Our findings provide novel insights into IL-33/mast cell-associated physiology and pathologies. PMID: 28259547
- Study showed that CLOCK can interact with RANBP9 and bind with mRNAs, demonstrating that CLOCK is involved in alternative splicing in spermatogenesis. These results reveal a novel mechanism for CLOCK in spermatogenesis. PMID: 29126923
- Disruption of CLOCK protein alters cortical circuits and leads to generation of focal epilepsy. PMID: 29024662
- Development of Mineralocorticoid receptor-mediated cardiac inflammation and fibrosis is dependent on intact signaling by the circadian protein CLOCK. PMID: 28911177
- Clock protein role in proteasomal and autophagic BMAL1 degradation and glucose homeostasis PMID: 26228022
- CLOCK transcription control of Wnt signaling promotes cell cycle progression in 3T3-L1 preadipocytes. PMID: 27194636
- results demonstrate that the core circadian gene Clock regulates bone formation via transcriptional control of 1,2,5(OH)2D3 receptor PDIA3 PMID: 27883226
- These results suggest that bone resorption and bone mass are regulated at a sophisticated level by osteoblastic Clock system through a mechanism relevant to the modulation of 1,25(OH)2 D3 -induced Rankl expression in osteoblasts. PMID: 27925286
- Abundance of CDH1 and TP63 proteins were significantly reduced in cultures transfected with shClock These data support how CLOCK plays a role in regulation of epithelial cell growth and differentiation in the mammary gland. PMID: 27707717
- These data establish the importance of circadian CLOCK-inhibin signaling in sepsis. PMID: 27520377
- our results indicate that Clock is a key regulator of AKT signalling for triggering cardiac hypertrophy in vivo. PMID: 28223222
- The findings indicate that the core circadian gene Clock may be essential during normal mouse embryonic stem cell (mESCs) differentiation by regulating mESCs proliferation, apoptosis and activity. PMID: 27664156
- CLOCK (a circadian clock component) is important for metabolic adaption of the heart to prolonged elevations in fatty acid availability. PMID: 26721420
- This study showed that the Clock delta-19 mutation reduces the average number of implantation sites and a significant lowering of the positions of implantation sites in the uterus, suggesting that the Clock mutation reduced the reproduction performance of the parents by affecting the implantation capacity via such as embryos' ability to implant. PMID: 27520292
- PI3K signaling plays a modulatory role in the regulation of the transcriptional rhythm of the Dbp gene by targeting BMAL1 and CLOCK. PMID: 27022680
- Findings indicate that CLOCK protein plays an important role in fertility and its knockdown leads to reduction in reproduction and increased miscarriage risk. PMID: 26390085
- This study demonstrated that ClockDelta19 mice exhibit rapid mood-cycling across the light/dark cycle. PMID: 25560763
- Clock gene expression in the suprachiasmatic nucleus in vivo is regulated by the circadian pacemaker and ultradian rhythms of unknown origin. PMID: 26194231
- In CLOCK-deficient fibroblasts, knockdown of Npas2 leads to arrhythmicity, suggesting that NPAS2 can compensate for loss of CLOCK in peripheral cells as well as in suprachiasmatic nucleus. PMID: 26895328
- SERPINA3K is involved in the Clock gene-mediated male fertility by regulating acrosin activity and provide the first evidence that SERPINA3K could be regulated by Clock gene via retinoic acid-related orphan receptor response elements. PMID: 26264441
- Circadian CLOCK Mediates Activation of Transforming Growth Factor-beta Signaling and Renal Fibrosis through Cyclooxygenase 2. PMID: 26458764
- Clock is a novel regulator of psoriasis-like skin inflammation in mice via direct modulation of IL-23R expression in gamma/delta+ T cells PMID: 26291684
- clock gene Rev-erbalpha and Wnt/beta-catenin signaling interact in the regulation of cell proliferation PMID: 25539035
- Insulin-phosphatidylinositol 3-kinase-Forkhead box class O3 (FOXO3) signaling is required for circadian rhythmicity in the liver via regulation of Clock. FoxO3 knockdown dampens circadian amplitude, an effect that is rescued by overexpression of Clock. PMID: 24856209
- Iso-mediated induction of Nfil3 in osteoblasts regulates the expression of Ptgs2 by driving the expression of circadian clock genes PMID: 24994935
- These data demonstrate a developmental role for circadian clock genes outside the suprachiasmatic nucleus, which may contribute mis-timed brain plasticity in associated mental disorders. PMID: 25801703
- The molecular clock governs rhythmic ncam1 alpha1-2-fucosylation in secondary olfactory neurons. PMID: 25384980
- Extinction of contextual fear was accelerated in mutant mice across the remaining retention sessions, as compared to WT mice, suggesting a role for Clock in extinction following aversive learning PMID: 24865659
- the suprachiasmatic nucleus clock is dispensable for the synchronization of peripheral clocks to the light-dark cycle PMID: 25063847
- identified novel regulators, endogenous receptors, in determining local clock activity, in addition to mediating the central control PMID: 25145629
- CLOCK phosphorylation may be essential for autonomous circadian oscillation in clock gene expression PMID: 25271155
- The results indicate that circadian clock gene expression is significantly modified in the suprachiasmatic nucleus and peripheral oscillators of aged mice. PMID: 24674978
- CLOCK and aryl hydrocarbon receptor nuclear translocator-like (BMAL)1 are required for hepatic circadian expression of sirtuin (SIRT)1. PMID: 24442997
- Hyperoxia-mediated lung injury is associated with alterations in CLOCK gene expression. PMID: 24696144
- The ClockDelta19 mutation appears to have no effect on the reinforcing properties of nicotine PMID: 24054990
- CLOCK-dependent direct transactivation through multiple E-boxes and indirect regulations polyphonically orchestrate dynamic circadian outputs. PMID: 24591654
- Clock plays a role in the ventral tegmental area as a negative regulator of ethanol intake and implicate the VTA dopamine system in this response. PMID: 23722243
- Data indicate that the protein expression levels of clock and apelin were significantly reduced when insulin-resistant adipocytes were treated with melatonin (Mel) and Neu-P11. PMID: 23963596
- this data suggest that clock genes in adipose tissue may play a role in metabolic adaptation to weight cycling. PMID: 24302006
- Together, our findings show that Clock is a positive regulator of ICAM-1, and promotes the adhesion of mononuclear cells to endothelial cells. PMID: 24333415
- These studies establish that circadian Clock activity is crucial in maintaining low plasma cholesterol levels and in reducing atherogenesis in mice. PMID: 24014832
- Functional CLOCK protein is essential to ensure the maintenance of mitochondrial content within muscle although the absence of a functional CLOCK protein does not impair the ability of animals to adapt to chronic exercise. PMID: 23429867
- Clock gene expression in round spermatids is essential for maintaining male reproductivity PMID: 23735500
- Findings indicate a clock-regulated mechanism of activation of the immune response against an enteric pathogen. PMID: 23716692
- the circadian expression of TF mRNA is regulated by clock molecules through a cell-autonomous mechanism via an E-box element located in the promoter region. PMID: 23291174
- Clock gene plays an important role in cocaine reinforcement and that decreased CLOCK function increases vulnerability for cocaine use. PMID: 22535308
- we show that 12-hour transcriptional rhythms (i.e., circadian "harmonics") are disrupted by Clock loss-of-function PMID: 22844252
顯示更多
收起更多
-
亞細胞定位:Nucleus. Cytoplasm. Cytoplasm, cytosol.
-
組織特異性:Expressed equally in brain, eye, testes, ovaries, liver, heart, lung, kidney. In the brain, expression is abundant in the suprachiasmatic nuclei (SCN), in the pyriform cortex, and in the hippocampus. Low expression throughout the rest of the brain. Expres
-
數(shù)據庫鏈接:
Most popular with customers
-
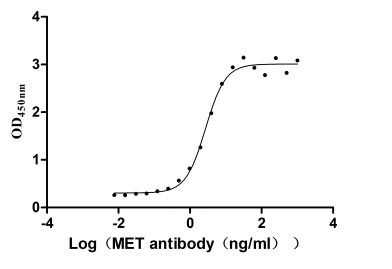
Recombinant Human Hepatocyte growth factor receptor (MET), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Human Mucin-16 (MUC16), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Human Receptor tyrosine-protein kinase erbB-2 (ERBB2), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Human Tumor necrosis factor ligand superfamily member 8 (TNFSF8), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Human Cannabinoid receptor 1 (CNR1)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Dog B-lymphocyte antigen CD20 (MS4A1)-VLPs (Active)
Express system: Mammalian cell
Species: Canis lupus familiaris (Dog) (Canis familiaris)
-
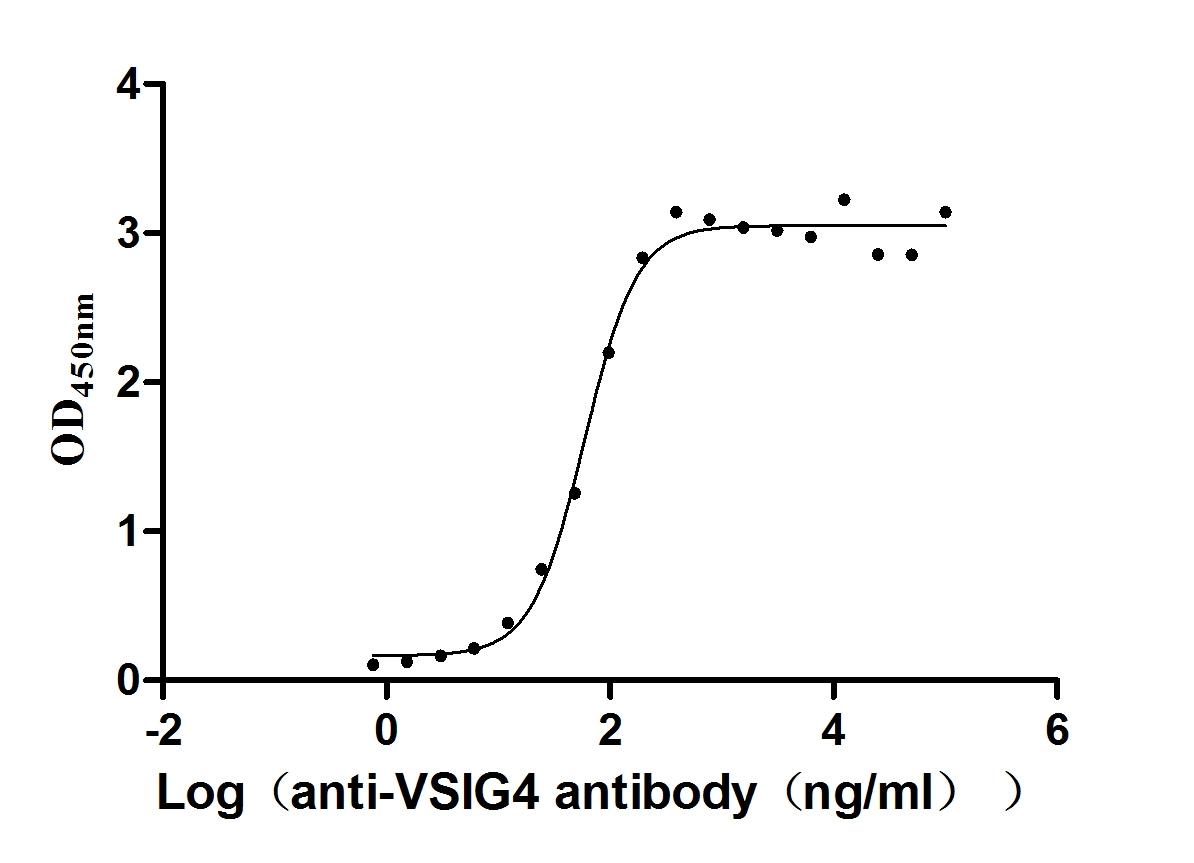
Recombinant Human V-set and immunoglobulin domain-containing protein 4 (VSIG4), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Macaca fascicularis Membrane spanning 4-domains A1 (MS4A1)-VLPs (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)