Recombinant Mouse Diacylglycerol kinase zeta (Dgkz), partial
-
中文名稱(chēng):小鼠Dgkz重組蛋白
-
貨號(hào):CSB-YP772140MO
-
規(guī)格:
-
來(lái)源:Yeast
-
其他:
-
中文名稱(chēng):小鼠Dgkz重組蛋白
-
貨號(hào):CSB-EP772140MO
-
規(guī)格:
-
來(lái)源:E.coli
-
其他:
-
中文名稱(chēng):小鼠Dgkz重組蛋白
-
貨號(hào):CSB-EP772140MO-B
-
規(guī)格:
-
來(lái)源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱(chēng):小鼠Dgkz重組蛋白
-
貨號(hào):CSB-BP772140MO
-
規(guī)格:
-
來(lái)源:Baculovirus
-
其他:
-
中文名稱(chēng):小鼠Dgkz重組蛋白
-
貨號(hào):CSB-MP772140MO
-
規(guī)格:
-
來(lái)源:Mammalian cell
-
其他:
產(chǎn)品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
別名:DgkzDiacylglycerol kinase zeta; DAG kinase zeta; EC 2.7.1.107; Diglyceride kinase zeta; DGK-zeta
-
種屬:Mus musculus (Mouse)
-
蛋白長(zhǎng)度:Partial
-
蛋白標(biāo)簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產(chǎn)品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復(fù)溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲(chǔ)存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質(zhì)期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項(xiàng):Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產(chǎn)品評(píng)價(jià)

靶點(diǎn)詳情
-
功能:Diacylglycerol kinase that converts diacylglycerol/DAG into phosphatidic acid/phosphatidate/PA and regulates the respective levels of these two bioactive lipids. Thereby, acts as a central switch between the signaling pathways activated by these second messengers with different cellular targets and opposite effects in numerous biological processes. Also plays an important role in the biosynthesis of complex lipids. Does not exhibit an acyl chain-dependent substrate specificity among diacylglycerol species. Can also phosphorylate 1-alkyl-2-acylglycerol in vitro but less efficiently and with a preference for alkylacylglycerols containing an arachidonoyl group. The biological processes it is involved in include T cell activation since it negatively regulates T-cell receptor signaling which is in part mediated by diacylglycerol. By generating phosphatidic acid, stimulates PIP5KIA activity which regulates actin polymerization. Through the same mechanism could also positively regulate insulin-induced translocation of SLC2A4 to the cell membrane. Regulates RASGRP1 activity.
-
基因功能參考文獻(xiàn):
- DGKzeta deficiency protects against peripheral insulin resistance and improves energy metabolism. PMID: 29066466
- DGKzeta prevents GLUT4 translocation in the absence of insulin. DGKzeta dissociated from IRS-1 by insulin stimulation enhances GLUT4 translocation through PIP5K1alpha activity PMID: 27739494
- Our studies identify a previously unrecognized DGKzeta contribution as a negative regulator of the crosstalk between phospholipase C-gamma- and phosphoinositide 3-kinase-regulated pathways. PMID: 28163304
- Our results demonstrate that, in addition to its known role in limiting canonical antigen-mediated activation of cytotoxic function, DGKzeta negatively regulates IL-2/IL-15-dependent expansion of innate-like cytotoxic CD8+ T cells. These studies add to the growing evidence that targeting DAG metabolism through pharmacological manipulation of DGKzeta could be an important, yet-unexplored area for cancer immunotherapy. PMID: 28438506
- these data suggest that the activation of DGKzeta downstream of antigen recognition provides a mechanism that ensures the activation of PA-dependent signaling as a direct result of the strength of TCR-dependent DAG mobilization. PMID: 27999176
- Diacylglycerol kinases alpha and zeta are up-regulated in cancer in cancer, and contribute towards tumor immune evasion and T cells clonal anergy. (Review) PMID: 27697466
- We propose that enzymes that negatively regulate distal activating receptor signaling pathways such as DGKzeta represent novel targets for augmenting the therapeutic potential of NK cells. PMID: 27342844
- Downregulation of DGKzeta might be one factor predisposing a person to osteolytic bone destruction in pathological conditions. PMID: 27312515
- this is the first report implicating M-CSF/DGKzeta/DAG axis as a critical regulator of bone homeostasis via its actions on OC differentiation and c-Fos expression. PMID: 25891971
- another possible involvement of DGKzeta in the regulation of secretion of the terminal tubule cells, as well as its functional significance of its nuclear localization in the submandibular ganglion cells is discussed PMID: 25952157
- This study demonstrated that Diacylglycerol Kinase zeta with Protein Kinase Calpha Is Required for Cerebellar Long-Term Depression PMID: 26586831
- The results suggest that DGKzeta cytoplasmic translocation in neurons under hypoxic stress is regulated by some mechanism which differs from that mediated by NAP1-like proteins. PMID: 24893663
- This study shows that DGKzeta knockdown facilitates degradation of IkappaB, followed by nuclear translocation of NF-kappaB p65 subunit. PMID: 25450975
- It plays a role in pathogenesis of various disorders in the central nervous system. PMID: 24599142
- DGKzeta regulates the development of natural Treg cells by limiting the extent of activation of the ERK and c-Rel signaling pathways. PMID: 24280042
- The zeta isoform of diacylglycerol kinase plays a predominant role in regulatory T cell development and TCR-mediated ras signaling. PMID: 24280043
- Data demonstrate that miR-34a is a negative regulator for DGKzeta and may play an important role in regulating T cell activation. PMID: 24147106
- Data indicate that in the absence of DGKzeta, the threshold for B cell antigen receptor (BCR) signaling, measured as activation of the Ras-extracellular signal-regulated kinase (ERK) pathway, was markedly reduced in mature follicular B cells. PMID: 24129701
- DGKzeta, but not PLD, plays an important role in mechanically induced increases in PA and mTOR signaling. PMID: 24302719
- The DGKzeta-deficient neurons do not succumb directly to apoptosis, although they are more vulnerable to excitotoxicity because of aberrant cell cycle reentry. PMID: 22516102
- Diacylglycerol kinase zeta serves as a sentinel to control p53 function both during normal homeostasis and in stress responses. PMID: 23606744
- Data indicate that combined deletion of dgkzeta and dgkalpha markedly enhances T-cell responses. PMID: 23576561
- As a regulator of Rac1 and RhoA activity, DGKzeta is a critical factor linking changes in lipid signaling to actin reorganization. PMID: 22918940
- DGK zeta is a crucial regulator of CXCL4-triggered T-cell arrest of surfaces presenting ICAM-1. PMID: 22546945
- These results suggest that DGKzeta regulates hippocampal long-term potentiation (LTP) and long-term depression (LTD) by promoting DAG-to-PA conversion, and establish that phospholipase C and protein kinase C lie upstream and downstream, respectively, of DGKzeta-dependent regulation of hippocampal LTP and LTD. PMID: 21069783
- DGKzeta inhibited ventricular tachyarrhythmias in a mouse model of heart failure and transient receptor potential channels participated in ventricular tachyarrhythmia induction in failing hearts. PMID: 21778596
- DAG kinases (DGKs) alpha and zeta, which terminate DAG-mediated signaling, synergistically inhibit TCR-induced mTOR activation by inhibiting the Ras-Mek1/2-Erk/12 pathway PMID: 21310925
- DGK-zeta export from nucleus to cytoplasm is regulated by a leucine-rich nuclear export signals through the exportin 1. PMID: 20023381
- Diacylglycerol kinase zeta functions as a physiological negative regulator of tcr signaling and t-cell activation PMID: 12883552
- DGK-zeta, syntrophin, and Rac1 form a regulated signaling complex that controls polarized outgrowth in neuronal cells PMID: 16055737
- DGKzeta may act in vivo as a downstream effector of pRB to regulate nuclear levels of diacylglycerol and phosphatidic acid PMID: 16286473
- Nuclear DGK-zeta might play some fundamental role during myogenic differentiation of C2C12 cells. PMID: 16897754
- These results are the first in vivo evidence that DGKzeta suppresses cardiac hypertrophy and fibrosis and prevents impaired left ventricular systolic function caused by pressure overload. PMID: 17251681
- DGKzeta promotes Toll-like receptors responses via a pathway involving inhibition of phosphatidylinositol 3-kinase PMID: 17371930
- Results suggest DGK-zeta and syntrophins play a role at multiple stages of the fusion process. PMID: 17410543
- DGK zeta blocks cardiac dysfunction and progression to lethal HF by activated G alpha q protein without detectable adverse effects in the in-vivo heart and suggest that DGK zeta is a novel therapeutic target for HF. PMID: 18219172
- Data show that ET-1-induced effects on Ca(2+) transients and cell shortening were abolished in diacylglycerol (DAG) kinase zeta-overexpressing mouse ventricular myocytes. PMID: 18275971
- DGKzeta localizes to the nucleus during interphase including G1, S, and G2 phases and is associated with chromatin although it dissociates from condensed chromatin during mitotic phase in NIH3T3 cells PMID: 18680142
- DGKs not only terminate diacylglycerol signaling but also initiate PA signaling in thymocytes to promote positive selection PMID: 18689679
- In DGKzeta-deficient fibroblasts PAK1 phosphorylation and Rac1-RhoGDI dissociation were attenuated, leading to reduced Rac1 activation after platelet-derived growth factor stimulation. PMID: 19211846
- Results show that diacylglycerol kinase zeta is targeted to excitatory synapses through its direct interaction with the postsynaptic PDZ scaffold PSD-95. PMID: 19229292
- Nuclear diacylglycerol kinase -zeta downregulated the expression of cyclin D1 and increased the expression of Btg2 protein, a transcriptional regulator of cyclin D1 with a strong anti-proliferative function. PMID: 19263516
- DGKz differentially regulates mast cell degranulation and cytokine production following FceRI stimulation PMID: 16717114
顯示更多
收起更多
-
亞細(xì)胞定位:Nucleus. Cytoplasm, cytosol. Cell membrane. Cell projection, lamellipodium.
-
蛋白家族:Eukaryotic diacylglycerol kinase family
-
數(shù)據(jù)庫(kù)鏈接:
Most popular with customers
-
-AC1.jpg)
Recombinant Human Angiopoietin-2 (ANGPT2) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
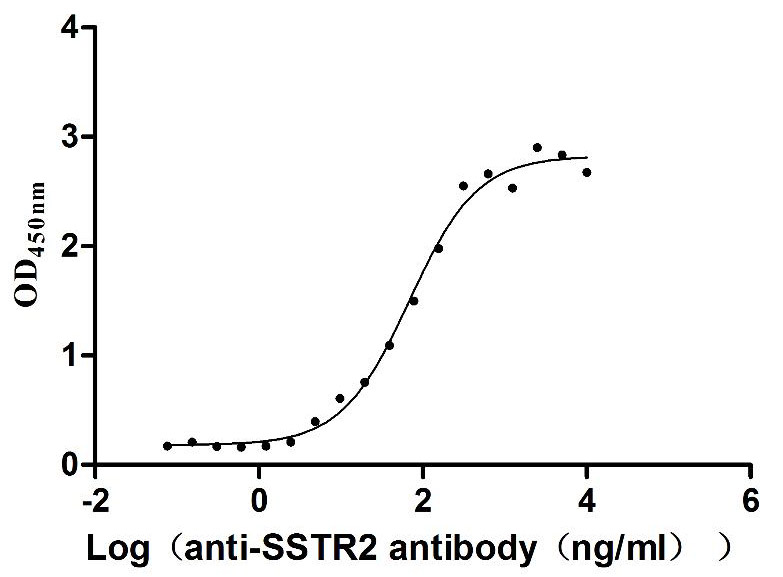
Recombinant Human Somatostatin receptor type 2 (SSTR2)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Dog B-lymphocyte antigen CD20 (MS4A1)-VLPs (Active)
Express system: Mammalian cell
Species: Canis lupus familiaris (Dog) (Canis familiaris)
-
-AC1.jpg)
Recombinant Human Claudin-6 (CLDN6)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Human Carcinoembryonic antigen-related cell adhesion molecule 8(CEACAM8) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Human C-C chemokine receptor type 6(CCR6)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)








