Recombinant Human Microprocessor complex subunit DGCR8 (DGCR8)
-
中文名稱:人DGCR8重組蛋白
-
貨號:CSB-YP845175HU
-
規(guī)格:
-
來源:Yeast
-
其他:
-
中文名稱:人DGCR8重組蛋白
-
貨號:CSB-EP845175HU
-
規(guī)格:
-
來源:E.coli
-
其他:
-
中文名稱:人DGCR8重組蛋白
-
貨號:CSB-EP845175HU-B
-
規(guī)格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:人DGCR8重組蛋白
-
貨號:CSB-BP845175HU
-
規(guī)格:
-
來源:Baculovirus
-
其他:
-
中文名稱:人DGCR8重組蛋白
-
貨號:CSB-MP845175HU
-
規(guī)格:
-
來源:Mammalian cell
-
其他:
產(chǎn)品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:DGCR8
-
Uniprot No.:
-
別名:GCRK6; C22orf12; D16H22S788E; D16Wis2; DGCR 8; Dgcr8; DGCR8 microprocessor complex subunit; DGCR8_HUMAN; DGCRK 6; DiGeorge syndrome critical region 8; DiGeorge syndrome critical region gene 8; Gy1; Microprocessor complex subunit DGCR8; pasha
-
種屬:Homo sapiens (Human)
-
蛋白長度:full length protein
-
表達區(qū)域:1-304
-
氨基酸序列METDESPSPLPCGPAGEAVMESRARPFQALPREQSPPPPLQTSSGAEVMDVGSGGDGQSE LPAEDPFNFYGASLLSKGSFSKGRLLIDPNCSGHSPRTARHAPAVRKFSPDLKLLKDVKI SVSFTESCRSKDRKVLYTGAERDVRAECGLLLSPVSGDVHACPFGGSVGDGVGIGGESAD KKDEENELDQEKRVEYAVLDELEDFTDNLELDEEGAGGFTAKAIVQRDRVDEEALNFPYE DDFDNDVDALLEEGLCAPKKRRTEEKYGGDSDHPSDGETSVQPMMTKIKTVLKSRGRPPT EPVL
-
蛋白標簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產(chǎn)品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質(zhì)期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產(chǎn)品評價

相關產(chǎn)品
靶點詳情
-
功能:Component of the microprocessor complex that acts as a RNA- and heme-binding protein that is involved in the initial step of microRNA (miRNA) biogenesis. Component of the microprocessor complex that is required to process primary miRNA transcripts (pri-miRNAs) to release precursor miRNA (pre-miRNA) in the nucleus. Within the microprocessor complex, DGCR8 function as a molecular anchor necessary for the recognition of pri-miRNA at dsRNA-ssRNA junction and directs DROSHA to cleave 11 bp away form the junction to release hairpin-shaped pre-miRNAs that are subsequently cut by the cytoplasmic DICER to generate mature miRNAs. The heme-bound DGCR8 dimer binds pri-miRNAs as a cooperative trimer (of dimers) and is active in triggering pri-miRNA cleavage, whereas the heme-free DGCR8 monomer binds pri-miRNAs as a dimer and is much less active. Both double-stranded and single-stranded regions of a pri-miRNA are required for its binding. Specifically recognizes and binds N6-methyladenosine (m6A)-containing pri-miRNAs, a modification required for pri-miRNAs processing. Involved in the silencing of embryonic stem cell self-renewal.
-
基因功能參考文獻:
- Primary microRNA transcripts (pri-miRs) are cleaved by Microprocessor, a complex containing the RNase Drosha and its partner protein, DGCR8. Although DGCR8 is known to bind heme, the molecular role of heme in pri-miR processing is unknown. Here we show that heme is critical for Microprocessor to process pri-miRs with high fidelity. PMID: 29170488
- Genotype AG in rs3757 DGCR8 exhibits protective effect, decreasing the risk of primary open angle glaucoma, while the homozygote GG is probably associated with increased risk of glaucoma. PMID: 29912490
- La protein is an important microprocessor component regulating miRNA processing efficiency by association with DGCR8 to regulate formation of the DGCR8-Drosha complex for miRNA processing. PMID: 29087193
- this study shows an unexpected function of DGCR8 in the repair of UV-induced DNA lesions that is independent of miRNA processing. PMID: 28380355
- BRG1 and SMARCAL1, members of the ATP-dependent chromatin remodelling family, are shown to co-regulate the transcription of DROSHA, DGCR8, and DICER in response to double-strand DNA breaks. PMID: 28716689
- Mechanistic dissection reveals that NEAT1 broadly interacts with the NONO-PSF heterodimer as well as many other RNA-binding proteins and that multiple RNA segments in NEAT1, including a 'pseudo pri-miRNA' near its 3' end, help attract the Drosha-DGCR8 Microprocessor. PMID: 28846091
- data suggest a model in which the bis-cysteine thiolate ligand environment of Fe(III) DGCR8 is necessary for establishing proper pri-miRNA binding and enabling processing activity. PMID: 27766492
- The rs417309 and rs1640299 polymorphisms of the DGCR8 gene as well as rs6877842 of the DROSHA gene might be associated with a risk of laryngeal cancer occurrence in the Polish population. PMID: 28155978
- studies support retention of DGCR8 cysteine coordination upon reduction, a conclusion distinct from those of previous studies of a different ferrous DGCR8 isoform PMID: 27546061
- Drosha and DGRC8 were significantly downregulated in healthy-appearing perilesional skin from hidradenitis suppurativa patients compared to healthy controls. PMID: 26917346
- Authors found that DENV4 infection exhibited the highest viral load 3 days post-infection. Dicer, Drosha, and DGCR8 showed reduced expression following DENV4 infection as compared with negative controls. PMID: 27173348
- Results demonstrated that DGCR8 is significantly upregulated in invasive ductal breast carcinoma, suggesting that increased expression of DGCR8 may play a fundamental role during the process of breast carcinogenesis. PMID: 26804549
- DGCR8 and Drosha assemble into a heterotrimeric complex on RNA, comprising two DGCR8 molecules and one Drosha molecule. PMID: 26683315
- Results show that DGCR8 forms an alternative complex with the RRP6-containing form of the exosome, acts as an adaptor to recruit the exosome to target structured RNAs, and the DGCR8/hRRP6 complex controls the stability of human telomerase RNA. PMID: 26687677
- We aimed to evaluate the expression of the major components of microRNA biogenesis machinery including Drosha, Dicer and DiGeorge syndrome critical region gene 8 (DGCR8) in multiple sclerosis patients PMID: 25439752
- These results reveal a new pathway in the DNA damage response wherein ABL-dependent tyrosine phosphorylation of DGCR8 stimulates the processing of selective primary miRNAs. PMID: 26126715
- DGCR8 functions as an oncogene in ovarian cancer, which is in part mediated by miR-27b. PMID: 25823356
- Together with a 23-amino acid peptide from DGCR8, DROSHA constitutes a minimal functional core. DROSHA serves as a "ruler" by measuring 11 bp from the basal ssRNA-dsRNA junction. DGCR8 interacts with the stem and apical elements through its dsRNA-binding domains and RNA-binding heme domain, respectively, allowing efficient and accurate processing. PMID: 26027739
- Data show decreased DGCR8 expression and miRNA dysregulation in individuals with 22q11.2 deletion syndrome PMID: 25084529
- These data show that hepatitis B virus proteins repress DGCR8 promoter activity by upregulating the expression of transcription factor YY1. PMID: 25427980
- in tumors with DGCR8 E518K and DROSHA exon 29 (miRNAPG-HS) mutations ... greater prevalence of tumors with blastemal predominant histology in patients with miRNAPG-HS and/or SIX1/2 Q177R mutations PMID: 25670082
- Recurrent mutations included a hotspot mutation (Q177R) in the homeo-domain of SIX1 and SIX2 in tumors with high proliferative potential (18.1% of blastemal cases); mutations in the DROSHA/DGCR8 microprocessor genes PMID: 25670083
- Study reveals a unique protein-RNA interaction central to pri-miRNA recognition. We propose a unifying model in which two DGCR8 dimers clamp a pri-miRNA hairpin using their Rheds. PMID: 24910438
- This study demonstrated for the first time that the DGCR8 mRNA expression level was up-regulated in colorectal carcinomas, suggesting its important role in pathobiology of colorectal carcinogenesis. PMID: 23775303
- multisite phosphorylation regulates DGCR8 protein stability, thereby raising microprocessor complex levels, changing the mature miRNA profile of the cell, and increasing cell proliferation and migration PMID: 24239349
- The Microprocessor complex of Drosha and DGCR8 proteins, which is responsible for the processing of the primary transcripts during the generation of microRNAs, destabilizes the mRNA of Aurora kinase B. PMID: 24589731
- A subset of senescence-associated miRNAs with the potential to target p21CIP1 is downregulated during DGCR8-mediated senescence. PMID: 23773483
- These data reveal a role for DeltaNp63 in the transcriptional regulation of DGCR8 to reprogram adult somatic cells into multipotent stem cells. PMID: 24449888
- It is a miRNA processing enzyme and altered in non-alcoholic fatty liver disease. PMID: 23663110
- specific RNA processing is likely facilitated by preformed DGCR8-Drosha heterodimers that can discriminate between authentic substrates and other hairpins PMID: 23893406
- RNase III enzyme Drosha and the double-stranded RNA-binding protein DGCR8 binds and regulates a large variety of cellular RNAs PMID: 23863141
- Single nucleotide polymorphisms in the DGCR8 3'-UTR that binds to miR-106b/miR-579 is associated with breast cancer. PMID: 23629745
- DGCR8, AGO1, AGO2, PACT, and TARBP1 expression levels were significantly higher in the epithelial skin cancer groups than the healthy controls (P > 0.05). PMID: 22025453
- DGCR8 and Drosha are targeted post-transcriptionally to chromosome 19 microRNA cluster pri-miRNAs as a preformed complex but dissociate separately. PMID: 22393237
- DGCR8-mediated cleavage of snoRNAs was independent of Drosha, suggesting the involvement of DGCR8 in cellular complexes with other endonucleases. Binding of DGCR8 to cassette exons is a new mechanism for regulation of alternatively spliced isoforms. PMID: 22796965
- DGCR8 is cleaved by caspases between Asp396 and Ser397 in HeLa cells. PMID: 22434730
- HDAC1 is an integral component of the Drosha/DGCR8 complex and enhances miRNA processing by increasing the affinity of DGCR8 to primary miRNA transcripts via deacetylation of critical lysine residues in the RNA-binding domains of DGCR8. PMID: 22222205
- This study demonstrates binding specificity of DGCR8 for ferric heme, provides direct biochemical evidence for ferric heme serving as an activator for miRNA maturation. PMID: 22308374
- analysis of the function of the DGCR8-heme interaction in microRNA maturation PMID: 21454614
- The mRNA for microprocessor component DGCR8 was found to be significantly upregulated in the dorsolateral prefrontal cortex and superior temporal gyrus in tissues from schizophrenic patients. PMID: 19721432
- The dimerization domain of DGCR8 is embedded in an independently folded heme-binding domain and directly contributes to association with heme. Heme-binding-deficient DGCR8 mutants have reduced pri-miRNA processing activity in vitro. PMID: 20506313
- SRY is a hybrid of DGCR8 and SOX3, and is regulated by the transcription factor CP2. PMID: 19902333
- a Microprocessor, containing the RNA binding protein Dgcr8 and RNase III enzyme Drosha, is responsible for processing primary microRNAs to precursor microRNAs PMID: 19759829
- Molecular cloning and expression analysis of DGCR8 PMID: 12705904
- DGCR8 may be an essential component of the primary miRNA processing complex, along with Drosha. PMID: 15574589
- DGCR8 is required for the maturation of miRNA primary transcripts. PMID: 15589161
- Thus, DGCR8 may function as the molecular anchor that measures the distance from the dsRNA-ssRNA junction PMID: 16751099
- Study show that the RNA-binding protein DiGeorge critical region-8 (DGCR8), which is essential for the first step of miRNA processing, is a heme-binding protein; the association with heme promotes dimerization of DGCR8. PMID: 17159994
- DGCR8 core recognizes pri-miRNA in two possible orientations. We propose a model for DGCR8's recognition of pri-miRNA PMID: 17704815
- DGCR8 locates at the nucleolus and small foci adjacent to splicing speckles in the nucleoplasm. PMID: 17765891
顯示更多
收起更多
-
亞細胞定位:Nucleus. Nucleus, nucleolus. Note=Colocalizes with nucleolin and DROSHA in the nucleolus. Mostly detected in the nucleolus as electron-dense granular patches around the fibrillar center (FC) and granular component (GC). Also detected in the nucleoplasm as small foci adjacent to splicing speckles near the chromatin structure. Localized with DROSHA in GW bodies (GWBs), also known as P-bodies (PubMed:17159994).
-
組織特異性:Ubiquitously expressed.
-
數(shù)據(jù)庫鏈接:
Most popular with customers
-

Recombinant Human Angiotensin-converting enzyme 2 (ACE2), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
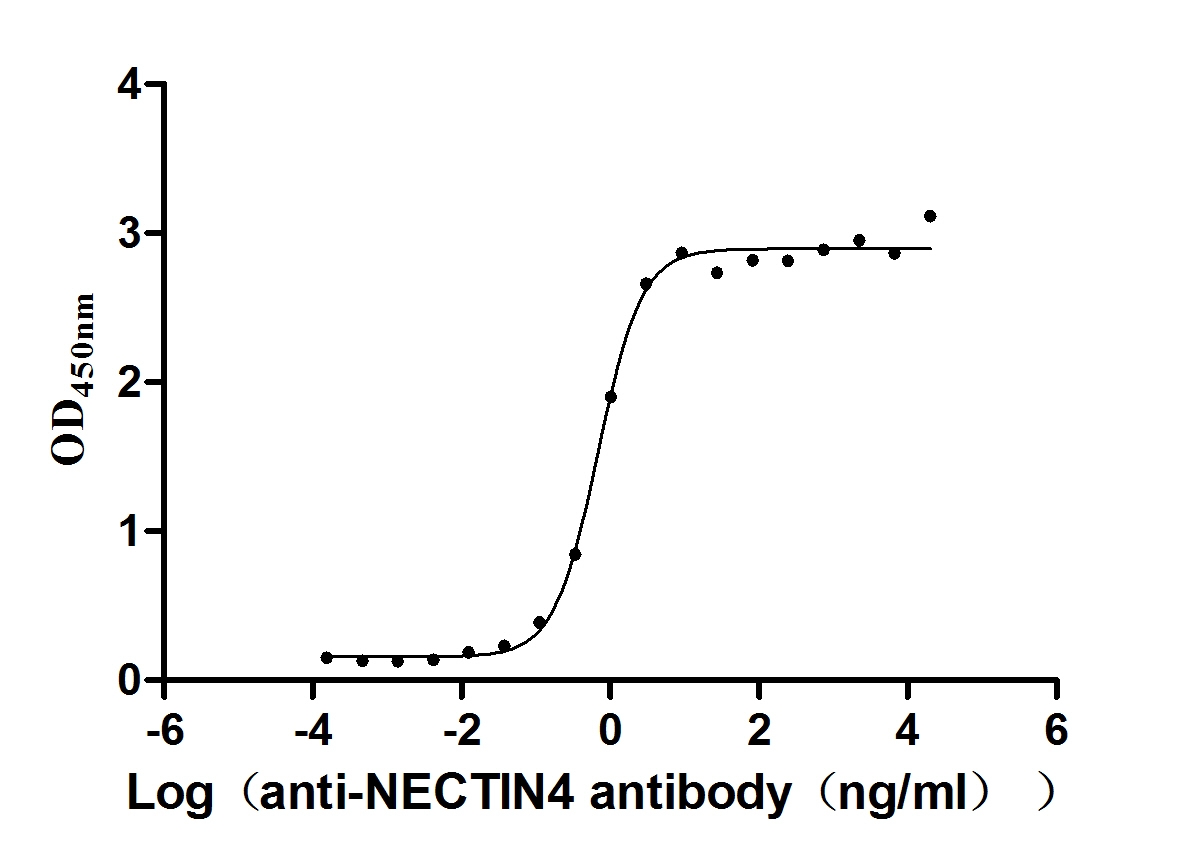
Recombinant Human Nectin-4 (NECTIN4), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
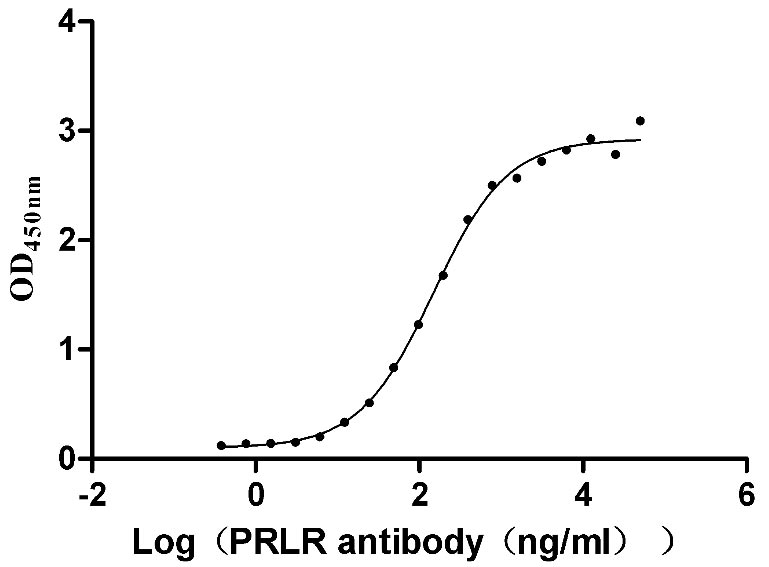
Recombinant Human Prolactin receptor (PRLR), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
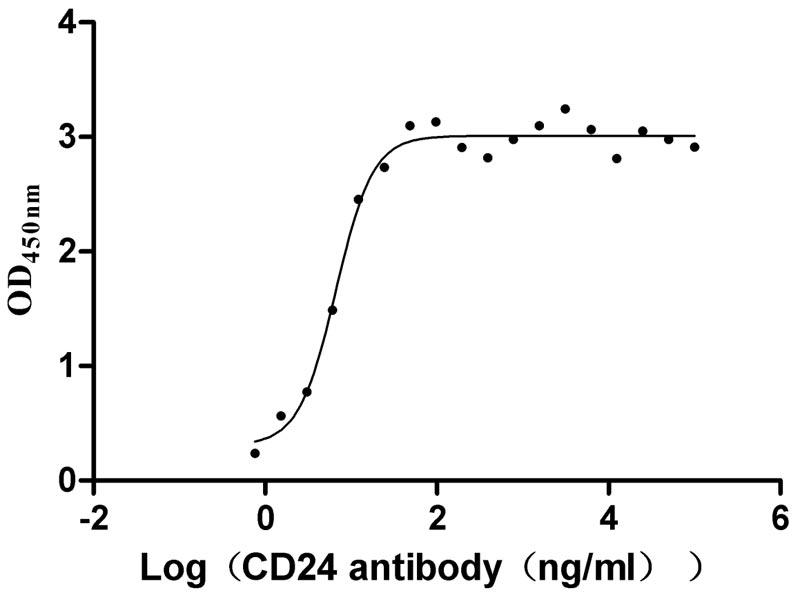
Recombinant Human Signal transducer CD24 (CD24)-Nanoparticle (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human G-protein coupled receptor family C group 5 member D (GPRC5D)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Mouse Tyrosine-protein kinase Mer (Mertk), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-

Recombinant Dog B-lymphocyte antigen CD20 (MS4A1)-VLPs (Active)
Express system: Mammalian cell
Species: Canis lupus familiaris (Dog) (Canis familiaris)
-

Recombinant Human Tomoregulin-2 (TMEFF2), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)