Recombinant Saccharomyces cerevisiae Cell division control protein 42 (CDC42)
-
中文名稱:Recombinant Saccharomyces cerevisiae Cell division control protein 42(CDC42),Yeast
-
貨號(hào):CSB-YP325635SVG
-
規(guī)格:
-
來源:Yeast
-
其他:
-
中文名稱:Recombinant Saccharomyces cerevisiae Cell division control protein 42(CDC42),Yeast
-
貨號(hào):CSB-EP325635SVG
-
規(guī)格:
-
來源:E.coli
-
其他:
-
中文名稱:Recombinant Saccharomyces cerevisiae Cell division control protein 42(CDC42),Yeast
-
貨號(hào):CSB-EP325635SVG-B
-
規(guī)格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:Recombinant Saccharomyces cerevisiae Cell division control protein 42(CDC42),Yeast
-
貨號(hào):CSB-BP325635SVG
-
規(guī)格:
-
來源:Baculovirus
-
其他:
-
中文名稱:Recombinant Saccharomyces cerevisiae Cell division control protein 42(CDC42),Yeast
-
貨號(hào):CSB-MP325635SVG
-
規(guī)格:
-
來源:Mammalian cell
-
其他:
產(chǎn)品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
別名:CDC42; SRO2; YLR229C; L8083.13Cell division control protein 42; Suppressor of RHO3 protein 2
-
種屬:Saccharomyces cerevisiae (strain ATCC 204508 / S288c) (Baker's yeast)
-
蛋白長(zhǎng)度:full length protein
-
表達(dá)區(qū)域:1-188
-
氨基酸序列MQTLKCVVVG DGAVGKTCLL ISYTTNQFPA DYVPTVFDNY AVTVMIGDEP YTLGLFDTAG QEDYDRLRPL SYPSTDVFLV CFSVISPPSF ENVKEKWFPE VHHHCPGVPC LVVGTQIDLR DDKVIIEKLQ RQRLRPITSE QGSRLARELK AVKYVECSAL TQRGLKNVFD EAIVAALEPP VIKKSKKC
-
蛋白標(biāo)簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產(chǎn)品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復(fù)溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲(chǔ)存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質(zhì)期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項(xiàng):Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產(chǎn)品評(píng)價(jià)

靶點(diǎn)詳情
-
功能:Involved in development of cell polarity during the cell division cycle, and essential for bud emergence. Affects signaling in the pheromone-response pathway through the STE20 protein kinase. Negatively regulated by the GTPase-activating proteins RGA1, BEM3, and BEM4.
-
基因功能參考文獻(xiàn):
- Using optogenetics, the authors probe yeast polarization and find that local recruitment of Cdc24 or Bem1 is sufficient to induce polarization by triggering self-sustaining Cdc42 activity. PMID: 28682236
- Here, the authors find that Bem1 directly augments the guanine exchange factor (GEF) activity of Cdc24. PMID: 28304276
- In Saccharomyces cerevisiae, the guanine nucleotide dissociation inhibitor (GDI) Rdi1 recycles Cdc42 through the cytoplasm. Loss of Rdi1 slowed but did not eliminate Cdc42 accumulation at the front, suggesting the existence of other recycling pathways. PMID: 27476596
- We find that Kss1 also localizes to the sites of polarized growth and Bem3 interacts with Kss1 at the septin-ring assembly site. TD2 localization is largely mediated by its interaction with the polarisome component Epo1 via heterotypic coiled-coil interaction. This finding reveals a novel role for the polarisome in linking Bem3 to its functional target,Cdc42. PMID: 28064039
- Fus2p localizes Cdc42p to the flat zone of cell fusion to promote cell wall degradation during cell fusion. PMID: 29066609
- we performed domain analysis to identify domain(s) of Gic1 that are important for its boundary activity, and identified two minimum domains, which are located outside its Cdc42-binding domain. PMID: 27301280
- cells evolved highly robust symmetry-breaking mechanisms that rely on feedback amplification of local Cdc42 activity. In turn, local Cdc42 activity favors Cdc42 accumulation because active Cdc42 is poorly mobile relative to the inactive form. PMID: 26338468
- Cdc42 activation must be localized for successful polarity establishment. PMID: 26459595
- Secretory vesicles deliver Cdc42p to sites of polarized growth in S. cerevisiae. PMID: 24945395
- Data indicate that Nba1 functions as a molecular spatial memory that confers protection from recurrent Cdc42 polarization at the same site. Authors propose that cytokinesis remnants are marked with "negative polarity cues" that prevent Cdc42 reactivation. PMID: 25416945
- Bem4p is a new regulator of the filamentous-growth MAPK pathway and binds to general proteins, like Cdc42p PMID: 25384973
- we have used electron microscopy and cryo electron tomography to elucidate the structural basis of the Gic1-septin and Gic1-Cdc42-septin interaction PMID: 24286829
- findings show that septins recruited to the site of polarization by Cdc42-GTP inhibit Cdc42 activity in a negative feedback loop and this inhibition depends on Cdc42 GTPase-activating proteins; findings define a fundamental mechanism underlying eukaryotic cell fate differentiation PMID: 23906065
- Activation of Cdc42 during mitosis causes a defect in cell separation. PMID: 23878274
- modeling predicts different dynamics of Cdc42-GTP polarization when the landmark level and the initial level of Cdc42-GTP at the division site are perturbed by noise added in the model. PMID: 23437206
- Cdc42 oscillations in yeasts. PMID: 23211526
- A novel regulatory input to Bni1p acting through the Cdc42p effector, Gic2p. PMID: 22918946
- identify a point mutant in the Rho-insert domain of CDC42, called cdc42-138, which is specifically defective in cell fusion PMID: 22323294
- Data show that Lem3-Dnf1/Dnf2 complex regulates the polar Cdc42 dynamics and phosphatidylserine distribution. PMID: 22344035
- Phosphatidylserine--and presumably its polarization--are required for optimal Cdc42 targeting and activation during cell division and mating. PMID: 21964439
- a small region at the N terminus is necessary and sufficient to confer Rho3 localization and function onto Cdc42 PMID: 20823269
- polarity regulators including Cdc42p were still polarized in mutants defective in both actin cables and cortical actin patches, suggesting that the actin cytoskeleton does not play a major role in cortical assembly of polarity regulators in budding yeast PMID: 20147449
- analysis of the solution structure of a novel Cdc42 binding module of Bem1 and its interaction with Ste20 and Cdc42 PMID: 20410294
- Results define a novel sterol-dependent subreaction of vacuole priming that activates cycles of Cdc42p activity to facilitate membrane fusion. PMID: 20007700
- Studies indicate that once Cdc42 is polarized to the presumptive bud site, it organizes the actin cytoskeleton, and the polarized secretion to carry out the polarized cell growth. PMID: 20175747
- These results define unique activation mechanisms for Cdc42p and Rho1p, which may be linked to the vacuole membrane fusion mechanism. PMID: 20171953
- Data show that Exo70 has the biochemical properties expected of a direct effector for both Rho3 and Cdc42, and suggest that interaction with the Exo70 component of the exocyst is a key event in spatial regulation of exocytosis by Rho GTPases. PMID: 19955214
- Findings show that the exocytic signal involves PI delivery Mss4 by Sfh5, generation of PI(4,5)P(2), and PI(4,5)P(2)-dependent regulation of Cdc42 and the actin cytoskeleton. PMID: 19477927
- Results suggest that the Cdc42 effectors Ste20, Cla4, and Skm1 translocate into the nucleus, where they modulate expression of sterol uptake genes via Sut1, thereby controlling sterol homeostasis. PMID: 19793923
- Using quantitative imaging and mathematical modeling, study elucidates mechanism of dynamic recycling of Cdc42 that balance diffusion; Rdi1, mediates a fast recycling pathway, while actin patch-mediated endocytosis accounts for a slower one. PMID: 20059952
- Cdc42p membrane localization and clustering requer Rho guanine-nucleotide dissociation inhibitors PMID: 15108020
- The opposite effects of overexpressed Pxl1p on cdc42 and rho1 mutants suggest that the functions of Cdc42p and Rho1p may be coordinately regulated during budding and that Pxl1p may be involved in this coordination. PMID: 15215315
- The ability of Rho3 and Cdc42 to hydrolyze GTP is not required for their role in secretion. PMID: 16103227
- Msb3 and Msb4 interact with Cdc42 and polarisome components to regulate cell polarity PMID: 16166638
- Data describe the presence of traveling waves of activated Cdc42p which randomly explore the cell periphery and confer dynamic responsiveness on the robust initiation of cell polarization. PMID: 16198298
- functions as a possible integrator for the upstream signals of filamentous differentiation (from the filamentous growth MAPK pathway, the cAMP pathway and the Mec1/Rad53 checkpoint pathway). PMID: 16200521
- Mutant phenotypes and suppression data suggest that the Cdc42p effectors Gic1p and Gic2p, previously implicated in polarization of the actin cytoskeleton, also function in septin recruitment. PMID: 16371506
- Cdc42p GDP-GTP cycling is critical for efficient cell fusion. PMID: 16571678
- Data from these studies have facilitated the temporal and spatial modeling of Rdi1p-dependent extraction of Cdc42p from the plasma membrane during the cell cycle. PMID: 17220465
- Sec14p regulation of phosphoinositide levels affects cytokinesis at the level of the Cdc42p/Cla4p/Ste20p signaling cascade. PMID: 17601877
- actin polymerization is a subreaction of vacuole membrane fusion governed by Cdc42p signal transduction. PMID: 17726018
- analysis of Cdc42p GTPase activation by cyclin-dependent protein kinases in budding yeast PMID: 17853895
- cooperation between protein-protein and protein-membrane interactions is a prevalent mechanism during Cdc42-regulated signaling and perhaps for other dynamic localization events at the cell cortex PMID: 17914055
- Bem2p and Bem3p may function as global inhibitors of Cdc42p activation during G1, and their inactivation by the Cdc28p/Cln kinase contributes to site-specific activation of Cdc42p at bud emergence PMID: 17914457
- a redistribution of phospholipids to the inner leaflet of the plasma membrane triggers the dispersal of Cdc42p from the apical growth site, through activation of GAPs. PMID: 17981141
- These findings highlight the important interactions among PLD1, Cdc42p and Ste20p during polarized cell growth in S. cerevisiae. PMID: 18036176
- The interactions of Sec3 with phospholipids and Cdc42 play important roles in exocytosis and polarized cell growth. PMID: 18195105
- Cdc24p oligomerization regulates Cdc42p activation via its localization PMID: 18378681
- Yeast symmetry-breaking polarization involves a guanine nucleotide exchange factor-p21-activated kinase complex that binds GTP-Cdc42p via the p21-activated kinase and promotes local Cdc42p GTP-loading via the GEF. PMID: 19013066
- Whi3 is required for the apical localization of Cdc28-Cln1,2 complexes during bud growth and thereby, to promote the activation of Cdc42 and its effectors in the bud apex. PMID: 19440046
顯示更多
收起更多
-
亞細(xì)胞定位:Cell membrane; Lipid-anchor; Cytoplasmic side.
-
蛋白家族:Small GTPase superfamily, Rho family, CDC42 subfamily
-
數(shù)據(jù)庫(kù)鏈接:
KEGG: sce:YLR229C
STRING: 4932.YLR229C
Most popular with customers
-

Recombinant Human Neuropilin-1 (NRP1) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Macaca fascicularis Claudin (CLDN18)-VLPs (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-

Recombinant Human Claudin-9 (CLDN9)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Human Claudin-4 (CLDN4)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Macaca fascicularis CD44 antigen (CD44), partial (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-

Express system: Mammalian cell
Species: Homo sapiens (Human)
-
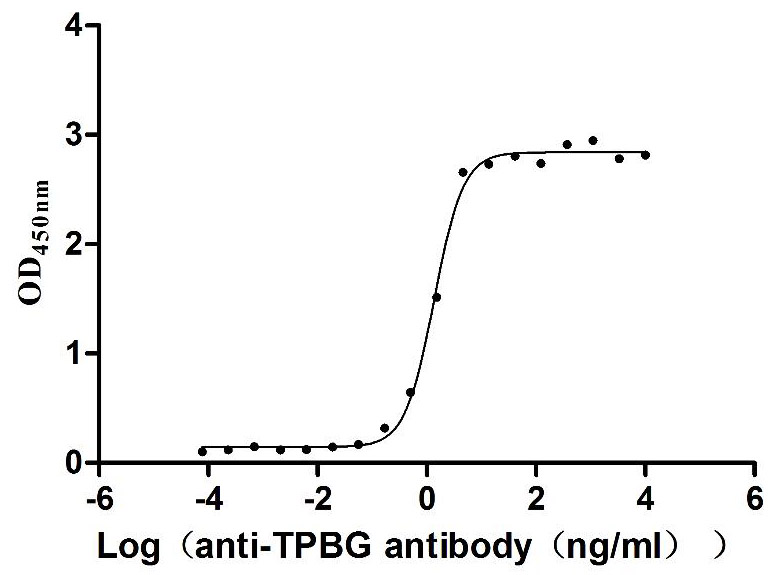
Recombinant Human Trophoblast glycoprotein (TPBG), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Human CD81 antigen (CD81), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)